ازدواج وقتی می تواند موفق و سازنده باشد که بدانید چه چیزی از آن می خواهید. چرا با این فرد به خصوص ازدواج می کنید و بعد از رفتن زیر یک سقف از او چه انتظاراتی دارید. در این صورت می توانید امیدوار باشید که با چشم باز راهی مسیر تاهل شده اید.
برای داشتن یک ازدواج موفق، باید قبل از ازدواج تکلیفتان را با این سوالات مشخص کنید:
سوالاتی که برای انتخاب یک همسر مناسب، باید به آن پاسخ داد
از علم عشق چه می دانید؟ آیا عشق در نگاه اول واقعی ست؟ بله. البته این عشقی رشد یافته نیست ، اما مغز عکس العملی سریع و پرشور به این محرک نشان خواهد داد. پس از اینکه شما کسی را که دوست دارید ملاقات کردید، مغز شما در کسری از ثانیه مواد شیمیایی مربوط به هرمون های عشق را آزاد می کند. نتایج مطالعه ای انجام گرفته در سال 2010 نشان داد که عشق واقعا مانند مواد مخدر عمل می کند. این جرقۀ عاشقانه همان احساس سرخوشی حاصل از مصرف کوکائین را ایجاد می کند .
دسته ای از دردهای جنسی که نیاز فوری به درمان دارند
دسته ای از دردهای جنسی که نیاز فوری به درمان دارند اگر به این درد جنسی مبتلا شوید بلافاصله باید درمان شوید و به پزشک مراجعه کنید و ... پریاپیسم به معنای نعوظ طولانیمدت، ناخواسته و مداوم آلت تناسلی مردان است که ناشی از تحریک یا برانگیختگی جنسی نیست و معمولا دردناک است.
این عارضه اگرچه شایع نیست اما نیاز به درمان طبی فوری دارد و در غیر این صورت به آسیب بافتی میانجامد و در نتیجه، فرد در آینده، هنگام برانگیختگی جنسی، نعوظ نخواهد داشت. پریاپیسم بیش از همه در پسران 5 تا 10 سال و مردان 20 تا 50 سال دیده میشود.....
ضربالاجل شش ماهه برای کسب مجوز سایتهای همسریابی همزمان با راهاندازی سامانه ثبتنام صدور مجوز برای مراکز مشاوره ازدواج شروع شد. به گفته معاون وزارت ورزش و جوانان، پس از گذشت شش ماه پلیس یا قوهقضاییه در صورت عدم تطبیق سایتهای همسریابی با مراکز مشاوره ازدواج، در مورد سایتهای همسریابی تصمیمگیری میکنند.
برنامه SkyGrabber یک دانلود کننده افلاین اینترنت ماهواره ای است و شما میتوانید اطلاعات ( movie, music, pictures) که توسط دیگر کاربران اینترنت استفاده و دانلود میشوند را دانلود و بر روی هارد دیسک خود ذخیره کنید. پس شما میتوانید فیلم های جدید بهترین موزیک ها وعکسها را دریافت نمایید. شما نیازی به کانکت شدن به اینترنت ندارید فقط شما باید تنظیمات دیش ماهواره را انجام داده و ماهواره مورد نظر را انتخاب نایید و سپس شروع به دانلود کنید. SkyGrabber یک برنامه بسیار جذاب و کاربر پسند میباشد و داری تنظیمات قابل انعطاف میباشد و اگر شما بخواهید تا بهترین نرم افزارها را به صورت رایگان دریافت نمایید اسکای گربر (SkyGrabber) میتواند بهترین انتخاب برای شما باشد. معمولا انواع مختلفی ارتباط با اینترنت می باشد مانند ( Dial-Up, ADSL, Leased Line) و غیره اینترنت ماهواره ای بیشتر در نقاطی که ناحیه های دور دست قرار دارند و استفاده از اینترنت بصورت دیگر امکان پذیر نیست و یا هزینه های هنگفت را در بر دارد.لینک دانلود ش
مشخصه های اسکای گربر : - شما به راحتی اطلاعات را بر اساس نیاز خود طبقه بندی و فیلتر انجام میدهید ( mp3, avi, mpg) - شما میتوانید حتی اطلاعات را براساس آدرسهای ای پی و مک (IP, MAC) فیلتر انجام دهید. - همزمان میتوانید از این برنامه استفاده کرده و به اینترنت وصل باشید - نشان دادن مراحل دانلود - پشتیبانی Kazza, Gnutella - به راحتی با بسته های ( TCP, GPE, IP, MPE) کار میکند - پشتیبانی از سیستم عامل های (Windows 95/98/Me/NT/2000/XP/Vista)
در عمل باکتریهایی که دارای خواص یکسانی باشند بندرت یافت میشوند، حتی باکتریهایی که از یک سلول منشا میگیرند ممکن است از نظر یک یا چند صفت با یکدیگر متفاوت باشند. این تفاوتها نتیجه تغییراتی است که به علت جهش ژنی یا موتاسیون در سلولهای باکتریایی پدید میآید. این باکتریهای تغییر یافته ، موتانت Mutant نامیده میشوند که از نظر بعضی از خواص نظیر ساختمان آنتی ژن ، حساسیت در مقابل آنتی بیوتیکها و ... با سایر باکتریهای مشابه اختلاف دارند.
سهولت تغییرپذیری در باکتریها مربوط به سرعت تقسیم آنهاست. زمان تقسیم یا مدت زمانی که برای تولید یک سلول جدید در باکتریها لازم است، حدود 2 دقیقه و در مورد انسان 20 سال است. مثلا یک سلول باکتری در مدت 18 ساعت 54 نسل بوجود میآورد. درحالیکه برای ایجاد همین تعداد نسل انسان بیش از 1000 سال زمان لازم است. پس جهش ژنی در باکتریها نسبت به موجودات عالی خیلی سریع و قابل ملاحظه است.
تفاوت یوکاریوتها با باکتریها
در کره خاکی تنها دو نوع سلول توسط کلیه ارگانیسمهای زنده تولید میشود. سلولهای پروکاریوت (یا هسته ابتدایی). در این گروه هسته ، فاقد غشا است و شامل کلیه باکتریهاست. پروکاریوتها شامل یوباکتریها (باکتریهای حقیقی) و آرکئی باکترها (باکتریهای قدیمی) است. اما گروه دیگر یوکاریوتها هستند که دارای غشای هسته و هسته حقیقی میباشند. اینگونه هسته در تمام ارگانیسمهای دیگر مانند Algae (جلبکها) Fungi (قارچها) ، پروتوزوئرها (protozoa) و گیاهان (Plant) و جانوران (Animals) یافت میشود. پاتوژنهای انسانی تنها در میان یوباکتریها یافت میشوند.
مشخصات سلول باکتری
اکثر باکتریها پوشش سلولی (cell envelope) تولید میکنند که شامل غشای پلاسمایی ، دیواره سلولی (cell wall) و پروتئینها و پلی ساکاریدهای تشکیل دهنده آن میباشد. بعضی از باکتریها کپسول یا لایه چسبنده تولید میکنند. فیلامانهای خارجی (فلاژل و پیلی) ممکن است در باکتریها بوجود آید. دیواره سلولی ، ساختمان سخت و مقاومی است که پروتوپلاست را احاطه کرده و آن را از آسیب فیزیکی و شرایط کاهش فشار اسمزی محیط خارج حفاظت میکند. معمولا به باکتری اجازه میدهد تا در برابر سطح وسیعی از شرایط محیطی ایستادگی کند پروتوپلاست از غشای سیتوپلاسمی و محتویات آن تشکیل شده است.
از نظر محتویات سلولی ، باکتریها سلولهای سادهای هستند. ساختمان اصلی سیتوپلاسم آنها شامل شبکه فیبریلی کروماتین مرکزی یا نوکلئوتید (Nucleoid) میباشد که توسط سیتوپلاسم بیشکل حاوی ریبوزومها احاطه شدهاست. اجسام انکلوزیون سیتوپلاسمی یا گرانولهای ذخیره انرژی ، بسته به گونههای باکتری ماهیت شیمیایی متفاوتی دارند و مقدار آنها به مرحله رشد و محیط بستگی دارد. بعضی از ساختمانهای سلولی از قبیل آندوسپورها فقط به تعداد کمی از باکتریها محدود میشوند.
طبقه بندی باکتریها
باکتریهای پست
این باکتریها تک یاختهای بوده و اگر کروی یا بیضوی باشند، کوکوس و اگر میلهای شکل یا دراز باشند، باسیل و اگر خمیده باشند ویبریون و چنانچه مارپیچی شکل و غیرقابل انعطاف باشند، اسپریل و اگر فنری و قابل انعطاف باشند، اسپیروکت نامیده میشوند.
باکتریهای عالی یا رشتهای
این باکتریها رشته مانند و اغلب غلافدار هستند و اغلب اوقات شاخههای حقیقی ایجاد کرده ، میسلیوم تشکیل میدهند و چون تشکیلات منشعب ایجاد میکنند، لذا اکتینومیست نامیده میشوند. بنابراین باکتریها از نظر شکل به 6 گروه گرد ، دراز ، خمیده ، مارپیچی ، فنری و منشعب تقسیم میشوند.
اجزای ساختمانی باکتریها
فلاژلها (Flagella)
فلاژلها ، فیلامانهای پروتئینی به طول و قطر یکنواخت میباشند و موجب تحرک شبیه به شنای سریع و مستقل اغلب باکتریها پاتوژنیک میگردند فلاژل در سه قسمت فیلامان ، قلاب و جسم پایه تشکیل شده است. پایه فلاژل در غشای پلاسمایی قرار گرفته است. لنگرگاه و تعداد فلاژل در باکتریها فرق خواهد کرد.
فیمبریاها
فیمبریاها که پیلی هم نامیده میشوند، فیبریلهای شبیه مو هستند به اندازه 0.004 تا 0.008 میکرون هستند. این ارگانل با میکروسکوپ الکترونی در سطح باکتریهای مختلف قابل رویت هستند. آنها مستقیمتر ، نازکتر و کوتاهتر از فلاژلها هستند. این رشتهها در غشای پلاسمایی سلول میکروبی لنگر میاندازد.
هسته باکتری
هسته سلول را میتوان بعد از رنگ آمیزی اختصاصی با میکروسکوپ نوری مشاهده کرد. در مقایسه با سلولهای عالی مواد ژنتیکی باکتریها و سایر سلولهای پست پراکنده ، ساده و بدون پوشش و کروموزوم حلقوی است غشای هسته وجود ندارد و کروموزوم به مزوزوم فرورفته در غشای سیتوپلاسمی چسبیده است. در سالهای اخیر پروتئینهای شبیه هیستون در باکتریها کشف شده است که احتمالا نقش مشابه هیستونها را در کروماتینهای سلولهای یوکاریوت ایفا میکنند.
سیتوپلاسم
بیش از 50 درصد پروتئین سلول در سیتوپلاسم قرار دارد و آنزیمهای متابولیسمی راههای گلیکولیز و بسیاری از آنزیمهای چرخه کربس ، انواع کاتالازها ، دهیدروژنازها ، و مواد حد واسط چرخه های متابولیکی در سیتوپلاسم وجود دارد. روابط اتمی ، یونی و الکترونی بین ترکیبهای مختلف سیتوپلاسمی با نظم خاص فعالیتهای حیاتی را ظاهر میسازد.
پوشش سلول (Cellenvelope)
کپسول و لعاب (Capsoles)
قدرت بیماریزایی پاتوژنها اغلب با تولید کپسول همراه است. باکتریهای کپسولدار در محیط جامد ، کلنیهای مخاطی (Mucoid) یا صاف (Smooth) میسازند. در مقابل باکتریهای فاقد کپسول کلنیهای خشن (Rough) دارند. اگر باکتری قدرت کپسولسازی خودش را از دست بدهد در مقابل قدرت ویرولانس (بیماریزایی) خود را از دست داده و در مقابل دستگاه ایمنی بدن میزبان تاب مقاومت نخواهد داشت.
دیواره سلولی
دیواره سلولی باکتریها بینهایت پیچیده است و لایه سفت و سختی را در اطراف باکتریها ایجاد میکند که سلول را از گسیختگی و متلاشی شدن در مقابل فشار اسمزی خارج سلول محافظت میکند. همچنین دیواره محل تجمع عوامل آنتی ژن میباشد که باکتریها را توسط این آنتی ژنها از هم تمیز میدهند. باکتریها با روش رنگآمیزی گرم (Gram stain) به دو دسته تقسیم میشوند.
گرچه هر دو گروه یعنی باکتریهای گرم مثبت و منفی دارای دیواره میباشند ولی فرق بین این دو گروه مربوط به خواصی است که در ساختمان دیواره سلولی آنها وجود دارد. اساس ساختمان در دیواره سلولی باکتریهای گرم مثبت یک لایه ضخیمی است از پپتیدوگلیکان (Poptidoglycan) ، ولی در باکتریهای گرم منفی ضخامت آن به حداقل میرسد.
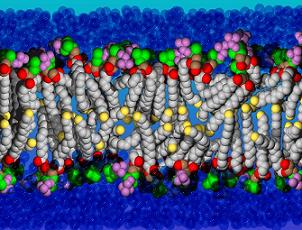
غشای سیتوپلاسمی
غشای سیتوپلاسمی غشای داخلی نیز نامیده میشود. غشای سیتوپلاسمی باکتریها مشخص بوده و از فسفو لیپید و پروتئین ساخته شده است. این غشا در پروکاریوتها از غشای سیتوپلاسمی در یوکاریوتها به علت نداشتن استرول متمایز میشود. چینخوردگیهای غشای سیتوپلاسمی به درون سلول ساختارهای ویژهای به نام مزوزوم ایجاد میکند که کروموزومهای باکتریها به مزوزومها متصل هستند. غشا همچنین به عنوان یک سد اسمزی برای سلول عمل میکند و دارای سیتوپلاسم انتقال دهنده برای مواد محلول است و انتقال تولیدات سلولی را در مقابل با محیط خارج سلولی تنظیم میکنند.
تولیدمثل باکتری
باکتریها به روشهای تقسیم مستقیم ، آمیختگی ، قطعه قطعه شدن یا بوسیله کنیدی و همچنین جوانه زدن تکثیر مییابند. برخی باکتریها توانایی ایجاد هاگ درونی را دارند. هاگ سبب مقاومت باکتری در برابر عوامل نامساعد محیط میشود. هر باکتری فقط یک هاگ میسازد و از هر هاگ یک باکتری بوجود میآید.
پیشرفتهایی که در سده اخیر نصیب علم ژنتیک شده است، تا حدود زیادی مرهون مطالعه و بررسی وراثت در باکتریها است. امروزه ثابت شده است که مکانیسمها ژنتیکی در باکتریها از نظر واکنشهای شیمیایی مشابه یاختههای یوکاریوت است. پروکاریوتها موجودات ساده و مناسبی برای بررسیهای ژنتیکی هستند. زیرا در آنها تنها یک مولکول DNA در هر یاخته وجود دارد و این DNA دارای ساختار کروموزمی پیچیدهای نیست. استفاده از میکروارگانیسمها به عنوان ابزار مطالعه ژنتیکی دارای نقاط ضعفی نیز است.
اول آنکه کوچکی اندازه این موجودات بررسی ویژگیهای ظاهری هر یاخته را دشوار میسازد. دوم آنکه تولید مثل جنسی در این موجودات وجود ندارد و یا بطور ناقص دیده میشود. پس از اینکه ساختار مولکولی DNA که نخستین بار بوسیله واتسون و کریک معرفی و ارائه شد، نحوه بیوسنتز آن را نیز در یاخته مشخص کردند. در اواخر سالهای 1950 ، کریک اصل بنیادی را مطرح کرد. این اصل بیان کننده چگونگی انتقال اطلاعات ژنتیکی از مولکول DNA به RNA و ترجمه آن در پروتئینها است.
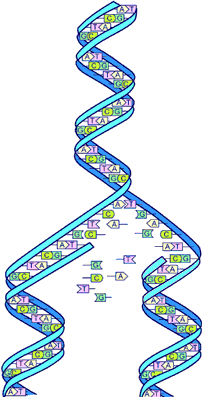
همانندسازی DNA
در مطالعات اولیه برای همانندسازی سه الگو مطرح شد که شامل الگوهای حفاظتی ، نیمه حفاظتی و پراکنده است. در الگوی حفاظتی از روی مارپیچ دو رشتهای DNA ، یک مولکول کامل DNA ساخته میشود. در الگوی نیمه حفاظتی ابتدا دو رشته DNA از هم باز شده و در مقابل هر یک از رشتهها ، رشته مکمل ساخته میشود. در الگوی پراکنده ابتدا مولکول DNA به قطعاتی تقسیم میگردد و هر یک از قطعه رشته مکمل خود را سنتز میکند. واتسون و کریک با پژوهشهای خود بر روی مولکول DNA ، الگوی نیمه حفاظتی را منطقی و تنها راه همانند سازی میدانستند. سپس مزلسون و استال با انجام آزمایشهای بسیار ظریف و مهم ، درستی چنین الگویی را به اثبات رساندند.
آزمایش مزلسون و استال
مزلسون و استال برای اثبات فرآیند همانند سازی آزمایشی انجام دادند که به شرح زیر میباشد. آنها ابتدا یاختههای باکتری اشرشیاکلی را در محیط کشت ویژهای که نیتروژن آن از نوع سنگین (N15) بود، برای زمان معین کشت دادند و سپس یاختهها را به محیط کشت عادی که نیتروژن آن از نوع سبک (N14) بود، انتقال دادند و در محدودههای زمانی معین از یاختههای نسلهای اول ، دوم و سوم حاصل از محیط کشت جدید ، نمونه برداری کرده و DNA آنها را به روشهای اختصاصی جدا ساختند. نمونههای DNA بر روی گرادیان (شیب) چگالی کلرور منیزیم سانتریفوژ شده و در این روش ترکیبات مختلف بر اساس چگالی آنها جدا سازی میشوند.
بدین ترتیب DNA واجد وزنهای متفاوت از یکدیگر جدا میشوند. DNA معمولی که N14 دارد (DNA سبک) به علت داشتن چگالی کمتر در بالای لوله قرار میگیرد. در حالی که مولکول DNA با (N15 سنگین) در محلی پایین تر از DNA سبک واقع میشود. DNA های واجد مقادیر متفاوت N15 و N14 نیز در بینابین این دو حد جای میگیرند.
با کشت یاختههای دارای DNA واجد نیتروژن سنگین در محیط کشت حاوی نیتروژن سبک مشاهده میشود که مولکول DNA ماهیت سبک - سنگین پیدا میکند. یعنی دو رشته DNA کاملا از هم باز شده و رشتههایی در تکمیل هر یک از دو رشته قبل ساخته میشود. این رشتههای جدید همگی دارای نیتروژن سبک (محیط کشت جدید) هستند. با ادامه کشت در نسلهای دوم و سوم ملاحظه میشود که از میزان DNA سبک - سنگین کم شده و به DNA سبک افزوده میشود.
نتیجه آزمایش مزلسون و استال
مزلسون و استال با چنین مشاهداتی نتیجه گرفتند که همانند سازی در مولکول DNA به طریق نیمه حفاظتی صورت میگیرد که مستلزم باز شدن دو رشته از هم و سنتز مولکول DNA جدید در مقابل هر رشته قدیم است. این پدیده به نام همانند سازی مشهور است.
آنزیمهای لازم در همانند سازی
آنزیمهای پلیمراز
آنزیمهایی هستند که پلیمر شدن زنجیرههای پلینوکلئوتیدی را کاتالیز میکنند. تا کنون سه نوع آنزیم پلیمراز به نامهای Ι و ΙΙ و ΙΙΙ جداسازی و مشخصات آنها ارائه شدهاند. از بین آنها آنزیم پلیمراز ΙΙΙ نقش اصلی را در سنتز DNA دارد. از خصوصیات مهم آن ، این است که منحصرا نوکلئوتیدها را در جهت '5 به '3 بهم متصل میکنند و در جهت عکس نمیتواند عمل کند. آنزیم پلیمراز ΙΙ نیز در مرحلهای از سنتز DNA وارد شده و سنتز را در جهت '3 به '5 پیش میبرد. و آنزیم پلیمراز I عمل ترمیم همانند سازی را انجام میدهد.
آنزیم هلیکاز
این آنزیم به مولکول DNA دو رشتهای متصل شده و با عمل خود موجب باز شدن دو رشته از یکدیگر میشود.
آنزیم لیگاز
در مرحلهای از سنتز DNA وارد عمل شده و دو رشته DNA را بهم پیوند میدهد.
آنزیم پریماز
آنزیمی است که در ساختن قطعه کوچک RNA پرایمر ، هنگام همانند سازی وارد عمل شده و نوکلئوتیدهایی از نوع اسید ریبونوکلئوتید را به یکدیگر متصل میکند. تعدادی پروتئینهای ویژه وجود دارند که پس از باز شدن دو رشته DNA از یکدیگر به محلهای باز شده متصل شده و مانع اتصال مجدد دو رشته به یکدیگر میشوند.
همانند سازی متوالی
در روی مولکول DNA نقاطی وجود دارند که همانند سازی از آنها آغاز میشود. این نقاط مبدا همانند سازی خوانده میشوند. در DNA باکتریها ، یک مبدا همانند سازی و در DNA موجودات عالی ، تعدادی زیادی از این مبدا وجود دارند. هنگام همانند سازی ابتدا آنزیم هلیکاز به مارپیچ دو رشتهای DNA متصل شده و پیچش DNA را در آن نقطه باز میکند. پرتئینهای DBP به ناحیه باز شده هجوم آورده و با اتصال به DNA تک رشتهای مانع از جفت شدن بعدی DNA میشوند.
ناحیهای را که هلیکاز به آن متصل میشود، چنگال همانند سازی مینامند. همانند سازی به صورت دو سویه است. آنزیم پلیمراز ΙΙΙ که اتصال نوکلئوتیدها را به یکدیگر به عهده دارد، فقط میتواند همانند سازی را در جهت 3 به 5 پیش ببرد. در این حالت دو رشته مولکول DNA در خلاف جهت یکدیگر هستند. در نتیحه رشتهای که در جهت '5 به '3 سنتز میشود، به راحتی سنتز DNA را آغاز کرده و پیش میبرد. این رشته به نام رشته راهنما معروف است. در همانند سازی این رشته را متوالی مینامند.
همانند سازی نامتوالی
در مولکول DNA رشتهای که '5 آزاد دارد، سنتز DNA طبق آنچه درباره رشته راهنما ذکر شد، انجام نمیگیرد. دلیل آن این است که آنزیم پلیمراز ΙΙΙ نمیتواند نوکلئوتیدها را در جهت 3 به 5 کاتالیز کند. لذا میبایست مکانیسم دیگری برای سنتز این رشته از DNA وجود داشته باشد. این رشته DNA به نام رشته عمل کننده یا پیرو معروف است. در این حالت ابتدا دو رشته DNA در فواصل معینی از یکدیگر باز شده و آنزیم پریماز در آن محل قرار میگیرد و با استفاده از ریبونوکلئوتیدها ، RNA کوچکی ساخته میشود که RNA پرایمر نام دارد.
انتهای 3 این RNA کوچک که از روی الگوی DNA ساخته شده است، میتواند به آنزیم پلیمراز III امکان دهد تا دزاکسی ریبونوکلئوتیدها را به انتهای آن متصل کند. لذا در این رشته از مولکول DNA قطعاتی از DNA سنتز میشوند که قطعات اوکازاکی نام دارد. (اوکازاکی نخستین کسی بود که این قطعات سنتز شده DNA را با میکروسکوپ الکترونی مشاهده کرد).
در این حالت آنزیم پلیمراز I وارد عمل شده و به ترتیب یکی یکی ریبونوکلئوتیدها را در جهت 5 به 3 برداشته و به جای آنها نوکلئوتیدهای از انواع دزاکسی جایگزین میکند تا این که قطعات همه از نوع دزاوکسی شوند. سپس انتهای قطعات ساخته شده بوسیله آنزیم لیگاز به هم متصل شده و یک رشته ممتد DNA حاصل میشود. اندازه هر قطعه اوکازاکی حدود 1000 تا 2000 نوکلئوتید است.
با استفاده از فنآوری DNA نوترکیب ، مطالعه ساختمان و عملکرد ژن بسیار آسان شده است و جداسازی یک ژن از یک کروموزوم بزرگ نیاز دارد به:
روشهایی برای برش و دوختن قطعات DNA
وجود ناقلین کوچک DNA که قادر به تکثیر خود بوده و ژنهایی در داخل آنها قرار داده شود.
روشهایی برای ارائه ناقل حاوی DNA خارجی به سلولی که در آن بتواند تکثیر یافته و کلنیهایی را ایجاد کند.
روشهایی برای شناسایی سلولهای حاوی DNA مورد نظر.
پیشرفتهای حاصل در این فنآوری ، در حال متحول نمودن بسیاری از دیدگاههای پزشکی ، کشاورزی و سایر صنایع میباشد.
پیشرفتهای حاصل از دهها سال کار هزاران دانشمند در زمینههای ژنتیک ، بیوشیمی ، بیولوژی سلول و شیمی فیزیک در آزمایشگاههای متعدد گرد هم آمدند تا فنآوریهایی برای تعیین موقعیت ، جداسازی ، آماده سازی و مطالعه قطعات DNA مشتق از کروموزومهای بسیار بزرگتر را ایجاد نمایند. تاکنون فنآوریهای کلون سازی DNA ، فرصتهای غیر قابل تصوری را برای تعیین هویت و مطالعه ژنهایی فراهم نمودهاند که تقریبا در هر فرآیند بیولوژیک شناخته شده ، نقش دارند. این روشهای جدید ، تحقیقات پایه ، کشاورزی ، پزشکی ، اکولوژی ، پزشکی قانونی و بسیاری از زمینههای دیگر را دگرگون کردهاند.
تخمیرهای میکروبی
تعدادی از محصولات مهم صنعتی بوسیله میکروارگانیزمها ساخته میشوند که از بین آنها ، آنتی بیوتیکها مهمترین گروه میباشند. بوسیله مهندسی ژنتیک میتوان میکروارگانیزمهایی ایجاد کرد که آنتی بیوتیک بیشتری تولید کنند و یا مشتقی از آنتی بیوتیک اولیه را بسازند.
واکسنهای ویروسی
واکسن مادهای است که میتواند سیستم ایمنی را بر علیه یک عامل عفونی تحریک کند. معمولا از ویروسهای کشته شده به عنوان واکسن استفاده میشود، ولی همواره یک خطر احتمالی وجود دارد که ویروس بطور کامل غیر فعال نشده باشد. از آنجایی که معمولا قسمت فعال و ایمنیزایی ویروس ، پروتئینهای پوشش آن هستند، میتوان پروتئینهای پوششی را به تنهایی و بدون قسمتهای دیگر تهیه کرد. برای این کار ژن مربوط به پروتئین پوششی را در یک باکتری و یا در یک ویروس غیر بیماریزا کلون میکنند و از آنها به عنوان واکسنهای بیخطر استفاده مینمایند.
تولید پروتئینهای خاص
تولید پروتئینهای خاص از نظر پزشکی و تجاری ارزش دارد. تولید تجاری پروتئینهای انسان از طریق استخراج از بافتها یا مایعات بدن غیر ممکن یا بسیار گران است. با کلون کردن ژنهای مربوط به این پروتئینها در باکتریها تولید تجاری این پروتئینها ، امکانپذیر میگردد.
حیوانات و گیاهان تغییر یافته
علاوه بر تولید محصولات ارزشمند بوسیله میکروبها ، از مهندسی ژنتیک میتوان به منظور ایجاد گیاهان و جانوران تغییر یافته استفاده کرد. به این گیاهان و جانوران بطور کلی تغییر یافته ژنتیکی (Trasgenetic) ، گفته میشود. تغییرات ژنی این موجودات ، مواردی چون تولید محصولات بیشتر ، تغییر کیفیت گوشت و سبزیجات و تولید پروتئینهای خاص که بوسیله باکتریها ، نمیتوان تولید کرد، را دربر میگیرد. این کار بطور کلی از طریق وارد کردن ژنهای نوترکیب در دوران جنینی به جانوران و در کشت بافت به گیاهان انجام میشود.
بیوتکنولوژی محیط زیست
باکتریها به دلیل تنوع متابولیزمی گسترده ، دارای یک خزانه ژنتیکی بسیار غنی میباشند. در بعضی موارد در این خزانه ژنهایی یافت میشوند که مواد آلوده کننده محیط زیست را تجزیه میکنند. ژنهای تجزیه بیولوژیکی بسیاری از مواد زاید فاضلابهای شهری و پسابهای صنعتی ، از باکتریهای موجود در طبیعت جدا شدهاند. از این ژنها میتوان برای کاهش آلودگیهای محیط زیست استفاده کرد.
مثالی از این کار ، ژنهای تجزیه کننده حشره کشهای کلردار ، مانند 5,4,2- تری کلروفنوکسی استیک اسید ، کلروبنزن ، نفتالین ، تولوئن ، آنیلین و هیدروکربنهای مختلف دیگر میباشد. ژنهای مورد نظر از باکتریهای پسدوموناس ، آلکالیژنس و تعدادی از باکتریهای دیگر جدا شده و در پلاسمیدهای مختلف وارد شده است. همچنین پلاسمیدهایی ایجاد شده است که ژنهای تجزیه کننده چند ماده مختلف را بطور همزمان بر روی خود دارند.
تنظیم ژنها و ژن درمانی
استفاده اولیه مهندسی ژنتیک در تولید محصولات مفید صنعتی و یا بهبود تولید بود، ولی مطالعات اخیر بر روی کنترل ژنهای خاص بنا شده است. امروزه قسمت اعظم تحقیقات پایه در مهندسی ژنتیک بر روی Antisense RNA که نقش مهمی در تنظیم ژنتیکی بیان ژنها به عهده دارد، پایه گذاری شده است. همچنین مطالعات گستردهای بر روی امکان درمان بیماریهای ژنتیکی از طریق وارد کردن ژن سالم یعنی ژن درمانی در حال انجام است.
تولید پروتئینها و هورمونهای کاربردی
یکی از کاربردهای عملی اولیه مهندسی ژنتیک تولید پروتئینهای مورد نظر بوسیله میکروارگانیزمهای سریعالرشد و تولید ارزان قیمت این پروتئینها بود. بسیاری از پروتئینها و پپتیدهای پستانداران ارزش دارویی زیاد دارند، ولی معمولا در مقادیر بسیار ناچیزی در بافتهای طبیعی وجود دارند و استخراج آنها مقرون به صرفه نمیباشد. این پروتئینها را میتوان به راحتی در میکروارگانیزمها تولید کرد.
تولید هورمونها
بسیاری از هورمونها ، پپتیدها و یا پروتئینهای کوچک هستند. این هورمونها در کنترل متابولیزم بدن پستاندارن و مخصوصا انسان استفادههای خاص و مهمی دارند. یکی از مثالهای این تولیدات ، تولید هورمون انسولین میباشد. هورمون انسولین انسانی اولین داروی تولید شده بوسیله مهندسی ژنتیک بود که مصرف عمومی پیدا کرد. انسولین هورمونی است که بوسیله غده لوزوالمعده ترشح میشود و کمبود آن باعث بیماری دیابت میگردد.
بیماری دیابت گریبانگیر میلیونها نفر در سراسر جهان است که روش استاندارد درمان آن ، تزریق منظم انسولین است. چون انسولین پستانداران مختلف تقریبا مشابه میباشد، در ابتدا از انسولین جدا شده از لوزوالمعده گاو و یا خوک استفاده میشد، ولی انسولین غیرانسانی به اندازه انسولین انسانی موثر نیست و هزینه خالص سازی نیز گران میباشد، لکن امروزه این هورمونها توسط مهندسی ژنتیک تولید میشوند.
لازم به ذکر است که تولید هورمونهایی مانند انسولین یک کار ساده مهندسی ژنتیک نیست که فقط شامل وارد کردن ژن مربوطه به داخل حامل و کلون کردن آن باشد، زیرا بسیاری از هورمونها فقط قطعات کوچکی از پلی پپتیدهای بزرگ تولید شده بوسیله ژنها میباشند.
چشم انداز
محصولات فنآوری DNA نوترکیب ، از پروتئینها تا موجودات مهندسی شده متفاوت میباشد. با این فنآوریها میتوان مقادیر زیاد پروتئینها را برای مقاصد تجارتی تولید نمود. از میکروارگانیزمها میتوان برای انجام کارهای اختصاصی استفاده نمود. با استفاده از مهندسی ژنتیک ، میتوان صفاتی را در گیاهان و جانوران ایجاد کرد که برای کشاورزی و پزشکی مفید باشند. بعضی از محصولات این فنآوری برای استفاده مورد تائید قرار گرفته و تعداد زیادی در حال تکامل هستند. در طی چند سال اخیر ، مهندسی ژنتیک از یک فنآوری وعده دهنده به یک صنعت چند بیلیون دلاری تبدیل شده و بیشتر رشد آن در صنعت دارویی بوده است.
کاربردهای مهندسی ژنتیک تقریبا نامحدود به نظر میرسد. این علم کاربردهای زیادی در علوم پایه و همچنین تولیدات صنعتی ، کشاورزی و علوم پزشکی دارد. در زمینه علوم پایه ، بررسیهایی مانند مکانیزمهای همانند سازی DNA و بیان ژنها در پروکاریوتها ، یوکاریوتها و ویروسها و همچنین چگونگی ساخته شدن و تغییرات پروتئینهای داخلی سلول و همچنین مکانیزم ایجاد سرطان از جمله کاربردهای مهندسی ژنتیک است. در زمینه کشاورزی که زمینه بسیاری از کاربردهای مهندسی ژنتیک بوده است، تولید گیاهان مقاوم به آفات گیاهی و خشکی ، تولید گیاهان پرمحصول و تولید گاوهای دارای شیر و گوشت بیشتر ، را میتوان نام برد. در زمینه کاربردهای انسانی ، تشخیص بیماریهای ارثی ، تولید انسولین انسانی ، تولید هورمون رشد انسان و ... را میتوان نام برد.
تاریخچه
اهمیت بعضی از اصول علمی ، در زمان کشف آنها مشخص نمیشود، بلکه پس از مدت زمانی که میگذرد ارزش آنها معلوم میشود. یکی از مثالهای روشن این مساله کشف ساختمان سه بعدی DNA بوسیله واتسون و کریک در سال 1953 بود. این ساختمان نسبتا ساده باعث شد تا دانشمندان سیستمهای مختلف ژنتیکی را بررسی کنند. اما مطلب به همین جا ، ختم نشد و دانشمندان مختلف سعی کردند که از این اطلاعات استفاده نمایند. هدف آنها نیز بیان سادهای داشت. آنها خواستند تا یک DNA را از یک موجود بگیرند و در موجود دیگر وارد نمایند تا اثرات آن ژن در موجود ثانویه بروز کند.
این علم نوین که به تدریج جای خود را در بین علوم دیگر پیدا کرد، با عناوین چون زیست مولکولی ، مهندسی ژنتیک و نهایتا DNA نوترکیب (Recombinant DNA) نامیده میشود. مثالی معروف از کارهای مهندسی ژنتیک تولید یک نوع باکتری اشرشیاکلی (E.Coli) است که قادر است انسولین انسانی بسازد. یا تولید گیاهان مقاوم به شوری و خشکی.
مراحل مهندسی ژنتیک
انتخاب ژن مورد نظر
جداسازی ژن مورد نظر
وارد کردن ژن مورد نظر در حامل
تکثیر ژن در میزبان مناسب
انتقال حامل ژن به سلول هدف
تکثیر سلول هدف
تولید انبوه محصول یا ایجاد صفت مورد نظر
تولید DNA نوترکیب با استفاده از آنزیمهای محدودالاثر(Restriction)
گروهی از آنزیم های محدودالاثر هنگام برش ، توالیهای مورد شناساییشان را بطور نامتقارن میشکنند، در نتیجه در انتهای قطعات DNA حاصله رشتههای تکی با حدود 4 نوکلئوتید بوجود میآید که به این انتهای تک رشتهای ، انتهای چسبنده (Sticky end) میگویند. یکی از آنزیمها ECORI نام دارد که باعث ایجاد قطعاتی میشود که در انتهای خود ، چسبنده میباشند.
حال فرض کنید که دو قطعه متفاوت DNA بوسیله یک آنزیم محدودالاثر یکسانی برش داده شدهاند، اگر قطعات حاصل از این برش با هم مخلوط شوند و شرایط مناسب فراهم شود انتهاهای چسبناک که مکمل هم میباشند بهم متصل میشوند. سپس بوسیله آنزیم DNA لیگاز این رشتهها به صورت کووالانسی بهم متصل میشوند.
هدف اصلی برش DNA در مهندسی ژنتیک ، اتصال دو قطعه DNA به یکدیگر میباشد. ولی هنگام اتصال قطعات DNA ممکن است بجای اینکه قطعات DNA بهم متصل شوند، دو سر یک مولکول DNA بار دیگر بهم بچسبند و در نتیجه نوترکیب صورت نگیرد. برای جلوگیری از این کار از آنزیم فسفاتاز قلیایی استفاده میکنند. به این صورت که پس از برش دادن حامل بوسیله آنزیم محدودالاثر فسفاتاز را به محیط واکنش میافزایند و در نتیجه فسفات انتهای 5 مولکول DNA در هر دو طرف جدا میشود و امکان اتصال دو سر مولکول حامل ، بدون DNA تازه ، به یکدیگر از بین میرود.
سیستمهای کلون کردن ژن
کلون کردن یک ژن خاص مهمترین مرحله مهندسی ژنتیک است. هدف از کلون کردن ژن به دست آوردن مقادیر زیادی از ژنهای خاص به صورت خالص میباشد. هدف اصلی کلون کردن ژن ، انتقال ژن مورد نظر از داخل یک ژنوم بزرگ و پیچیده به داخل یک حامل ساده و کوچک تکثیر آن است.
مراحل کلون کردن ژن
جداسازی و قطعه قطعه کردن منبع DNA: منبع DNA میتواند، ژنوم کامل یک موجود باشد که در این صورت، باید آن را بوسیله آنزیم محدودالاثر برش داد و قطعات حاصله را برای کلون کردن بکار برد.
اتصال به یک حامل کلون (Cloning Vector): حاملهای کلون ، قطعات ژنتیکی کوچکی هستند که بطور مستقل توانایی تکثیر دارند و دارای محل برش بوسیله آنزیمهای محدودالاثر میباشند، ولی این برش نباید در محل همانند سازی این حاملها باشد.
ورود به داخل میزبان:DNA نوترکیب حاصل به روشهای مختلف وارد باکتری یا میزبان مورد نظر میشود.
شناسایی و جداسازی کلون حاوی ژن مورد نظر: این مرحله شامل جداسازی میزبانهایی است که ژن مورد نظر بوسیله حامل وارد آنها شده و به نحو موثر بیان میشود.
تولید تعداد زیاد سلولها و یا باکتریهای حاوی ژن: این کار به منظور جداسازی و بررسی ژن مورد نظر ، انجام میگیرد.
حاملهای کلون (Cloning Vector)
پلاسمیدها
قطعات DNA حلقوی هستند. که در داخل سیتوپلاسم باکتریها و جدا از کروموزوم آنها قرار دارند و بطور مستقل تکثیر میشوند. پلاسمیدها ، خصوصیات مفیدی برای استفاده به عنوان حامل دارند مانند: اندازه کوچک ، DNA حلقوی ، همانند سازی مستقل ، تکثیر زیاد و شاخصهای مفید دیگر مانند دارا بودن ژنهای مقاومت به آنتی بیوتیک که جداسازی کلنیهای حاوی پلاسمید را راحتتر میکند.
باکتریوفاژها (ویروس باکتری)
ویروسها به خاطر داشتن پروتئینهای خاص ، نفوذ بسیار موثر و اختصاصی را به داخل سلولهای میزبان انجام میدهند.
بعضی ویروسها در قسمتی از چرخه تکثیر خود ، نفوذ پایداری به داخل ژنوم میزبان دارند که این باعث پایداری بیان ژن در داخل سلول میزبان میشود.
ویروسها دارای پروموتورهای خاصی هستند که بوسیله سلولهای میزبانی شناخته میشوند و این باعث بیان مناسب ژنهای کلون شده میشود.
کازمیدها (Cosmids)
کازمیدها در حقیقت قطعات حاصل از دو انتهای ژنوم از دو انتهای ژنوم باکتریوفاژها لامبدا قرار بگیرند و در نتیجه وارد سلول Ecoli (باکتری اشرشیاکلی)شوند. در داخل سلول E.Coli این DNA به صورت حلقوی در آمده و مانند یک پلاسمید عمل می کند.
فاسمیدها
یکی دیگر از حاملهای DNA نوترکیب هستند که ترکیبی از ژنوم باکتریوفاژ و پلاسمیدها هستند.
انتخاب میزبان مناسب
میزبان مورد نظر باید خصوصیاتی از قبیل پایداری ژنتیکی ، ژنوم کاملا شناخته شده مشخصات فیزیولوژیک معلوم ، توانایی پذیرش DNA خارجی ، داشتن یک شاخص خاص برای شناسایی در مواقع لزوم و ... را داشته باشد. یکی از شناخته شده ترین میزبانهای مورد استفاده باکتری E.Coli است. هنگام انجام کارهای ژنتیکی باید با مطالعاتی کافی یک سیستم حامل میزبان مناسب را انتخاب کرد و بکار برد. باسیلوس سوبتلیس (B.Subtilis) در مواردی که هدف از کلون کردن تولید یک پروتئین خالص میباشد، بر E.Coli ترجیح دارد. زیرا خصوصیات تخمیری این باکتری برای تولیدات صنعتی مناسب تر است.
روشهای وارد کردن حاملها به داخل میزبان
ویروسها و باکتریوفاژها
برای ویروسها و باکتریوفاژها و همچنین DNA نوترکیب که در داخل کپسید ویروس ها قرار گرفتهاند (کاسمیدها) روش ورود واضح است و همانند ورود معمولی ویروس ها در سلول های میزبان است.
ترانسفورماسیون: برای این کار DNA نوترکیب را با باکتری مجاور میکنند. این روش یکی از متداولترین روشهای انتقال است.
الکتروپوریشن: در این روش قطعات DNA را در یک محیط دارای بار الکتریکی در مجاورت سلولها قرار میدهند. بار الکتریکی باعث ایجاد منافذ ریز در غشای سیتوپلاسمی میشود که این خود باعث تسهیل ورود قطعات DNA به داخل سلول میگردد.
تفنگ ذرهای یا تفنگ اسید نوکلئیک: در این روش دقیقا تنگی در مقیاس میکروسکوپی وجود دارد که گلوله آن قطعات DNA میباشد و DNA را به داخل سلول ، شلیک میکند.
انتخاب کلونهای تغییر یافته
پس از اینکه DNA نوترکیب ساخته شد و در داخل باکتری میزبان ، انتقال داده شد. حال نوبت به انتخاب کلونهای باکتریایی میرسد که DNA نوترکیب مورد نظر به داخل آن انتقال یافته و به نحو موثری در داخل آن بیان شود. 3 خصوصیت در بین حاملین مشترک است. قدرت تکثیر در میزبان ، محل ورود ژن خارجی و یک شاخص انتخابی.
شاخصهای انتخابی موجود بر روی حاملها
مقاومت به آنتی بیوتیکها
مقاومت به آنتی بیوتیکها معمولا یا بوسیله آنزیم هایی ایجاد میشود که باعث غیر فعال شدن آنتی بیوتیکها میشوند و یا با سنتز پروتئینهایی است که به روشهای مختلف باعث ممانعت از اثر آنتی بیوتیکها میشوند. هر دو نوع مکانیزم مقاومت فوق بوسیله قطعات ژنتیکی ، کنترل میشوند. این قطعات ژنتیکی را میتوان در حاملها وارد کرد و از آنها به عنوان شاخصهای انتقال موثر استفاده کرد.
نیازهای متابولیزمی
نیازهای متابولیزمی طیف وسیعی از مواد مختلف را شامل میشود. برای این کار از گونههای خاص از میزبان استفاده میشود که تونایی ساختن یک ماده متابولیزمی ضروری از دست دادهاند، در نتیجه این باکتریها بر روی محیطهای بدون این ماده متابولیزمی رشد ، نخواهد کرد. برای مثال اگر یک باکتری توانایی تولید اسید امینه لوسین را نداشته باشد. بر روی محیط فاقد لوسین رشد نخواهد کرد.
حال اگر ما از حاملی استفاده کنیم که حاوی ژن سنتز لوسین باشد، باکتریهای میزبان حاوی این حاملها بر روی محیط فاقد لوسین رشد خواهند کرد. پس از اینکه کلنیهای حاوی ژن نوترکیب انتخاب و جدا شدند، این کلنیها را به میزان دلخواه تکثیر میدهند و سپس ژن تکثیر شده را برای بررسیهای بعدی استخراج کرده قرار میدهند.
حاملهای بیان ژن (Expression Vector)
یک حامل بیان ژن حاصل است که نه تنها میتوان از آن به عنوان حامل کلون استفاده کرد. بلکه این حامل دارای کی توالی تنظیمی میباشد که باعث میشود که بیان ژن مورد نظر تحت کنترل مهندسی ژنتیک قرار گیرد. یک حامل بیان ژن خوب باید دارای مشخصات زیرا باشد. هر چه قدر تعداد نسخههای یک ژن بیشتر باشد، میزان بیان آنها بیشتر خواهد بود. پلاسمیدها از این نظر مناسب هستند. قدرت آغازگری آن خوب باشد. الگوی خواندن آن مناسب باشد. بطور کلی وظیفه مهندسی ژنتیک ایجاد یک حامل مناسب است که بتوانند بطور موثری به داخل میزبان وارد شود به تعداد همانند سازی کند بطور موثر نسخه برداری شود - بطور موثر ترجمه شود.


.jpg)












 چشماتو از گوشه بکش بعد نگاه کن
چشماتو از گوشه بکش بعد نگاه کن